 Кровь выполняет многочисленные функции только тогда, когда циркулирует сердечно-сосудистой системой. В организме есть два замкнутых круга кровообращения-малый (легочный) и большой (системное). Подключенные они последовательно.
Кровь выполняет многочисленные функции только тогда, когда циркулирует сердечно-сосудистой системой. В организме есть два замкнутых круга кровообращения-малый (легочный) и большой (системное). Подключенные они последовательно.
Большой круг кровообращения начинается из левого желудочка, который выбрасывает кровь в аорту. От аорты отходят многочисленные артерии, питающих отдельным органам. Вследствие ветвления артерий и артериол образуются капиллярные сосуды, которые обеспечивают обмен между кровью и клетками. При слиянии капилляров образуются венулы, которые составляют вены. Последние, объединившись в нижнюю и верхнюю полые вены приносят кровь в правое предсердие.
Малый круг кровообращения начинается из правого желудочка, который выбрасывает кровь в легочный ствол, и сосудами легких она оттекает через четыре легочные вены в левое предсердие.
Четырехкамерное сердце объединяет эти круги в единую систему кровообращения. Непрерывный кровоток возможен лишь при условии, что правое и левое отделение сердца работают синхронно и равномерно: оба желудочка при каждом сокращении должны выбрасывать в соответствующие
сосуды одинаковое количество крови.
В сосуды кровь поступает Гири сокращении желудочков (систола). При их расслаблены (диастола) все полости сердца заполняются кровью. Расположенные перед желудочками предсердия способствуют приема крови из сосудов при сокращении желудочков. Предсердия не только депонированные кровь притекает к сердцу, но и подают желудочков у.рази необходимости дополнительную порцию крови. Особое значение эта дополнительная «подкачка» приобретает при увеличении частоты сокращений сердца, когда диастолическое становится короче и уменьшается продолжительность заполнения полостей желудочков кровью. Примерно такую же функцию для предсердий выполняют их ушки.
Кардиомиоциты. Структурной основой сердца миокард. Он имеет массу 250-300 г. Толщина его расположение волокон и даже собственно волокна в различных отделах не одинаковы. Миокард предсердий тоньше, чем миокард желудочков, и имеет два слоя: циркулярный-и продольный. Циркулярные волокна окружают преимущественно сосуды, впадающие в предсердия и Гири сокращении перетискують их отверстия. Миокард желудочков, особенно левого, мощный и состоит из трех слоев. Наружный и внутренний слои имеют спиралевидную форму и являются общими для обоих желудочков. Средний слой состоит из циркуляторной расположенных мышечных волокон. Он изолирован в правом и левом желудочках. Волокна предсердий и желудочков прикрепляются к фиброзного кольца.
К нему крепятся и клапаны сердца. Диаметр кардиомиоцитов предсердий меньше, чем желудочков.
Кардиомиоциты имеют прямоугольную форму, длину - около 120 мкм и ширину - 17-20 мкм. Содержат все структуры, характерные для волокон поперечно скелетной мышцы: ядра, миофибриллы, митохондрии, пластинчатый комплекс, саркоплазматический ретикулум. Но саркоплазматического ретикулума (депо Са2 +) в нем меньше, чем в скелетных мышцах
Между отдельными кардиомиоцитами существуют многочисленные межмембранного контакты - нексус (вставные диски). Они обеспечивают электрические взаимодействия между клетками, образуя функциональный синцитий миокарда. Такая структура обеспечивает сокращения кардиомиоцитов со сравнительно большой скоростью и развивает достаточно мощные усилия для выбрасывания крови.
Проводящей системы сердца. Среди сократительного миокарда расположена система вдитин и отростков, которые выполняют функции генерации возбуждения и проведения его в кардиомиоцитов. Это и есть проводниковая система сердца. Кардиоциты этой системы сгруппированы в определенных участках миокарда в узлы. Они меньше по размеру, чем сократительные элементы предсердий и желудочков, соединяются в группы, имеющие большое количество взаемнопереплетених выростов. В их структуре более саркоплазмы и мало миофибрилл, слабо Т-система саркоплазматического ретикулума. По своим свойствам приближаются к эмбрионального миокарда; устойчивы к гипоксии, поскольку в них энергия образуется в основном за счет анаэробного гликолиза.
Проводниковая система состоит из синусного и атриовентрикулярного узлов, межузловых и межпредсердной коммуникаций, предсердно-желудочкового пучка (пучка Гиса), который делится на
правую и левую ножки, идущие по межжелудочковой перегородке в субэндокардиальных сетку миоциты сердечного ведущего, или волокон Пуркинье.
Синусовый (синусно-предсердный) узел находится у места впадения верхней и нижней полых вен в правое предсердие. Узел имеет эллипсоидную форму, длину 10-15 мм, ширину 4-5 мм, толщину 1,5 мм. Он состоит из «головки» и «хвостовой» части. «Головка» лежит субэпикардиально. От нее до ушка левого предсердия отходят пучок волокон, который был описан J. G. Bachmann (1916), задний мижвенозний пучок (К. F. Welckenbach, 1907) и задний межузловой пучок (Ch. Thorle, 1909). Эти пучки передают возбуждение атриовентрикулярного узла.
Атриовентрикулярный узел находится в толще межпредсердной перегородки на границе предсердий и желудочков. Размер узла -
7,5 X3, 5X1 мм.
Пучок Гиса имеет длину 12-40 мм, ширину 1-4 мм. Он начинается стволом от атриовентрикулярного узла и, пройдя через межжелудочковой перегородки, делится на две ножки, идущие по левому и правому сторонам межжелудочковой перегородки и переходят в волокна Пуркинье, которые непосредственно контактируют
с клетками сократительного миокарда.
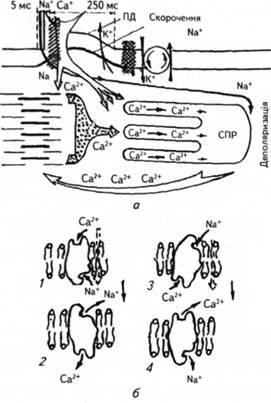
Електровозбудимость сердца. Ведущая и сократительная функции сердца выполняются за счет ионных перемещений, которые образуют электрические потенциалы. Развитие их зависит главным образом от концентрации свободных ионов внутри и снаружи клеточных мембран и их проницаемости. Обмен ионов между внешним и внутренним средами клетки ВЧ первую очередь зависит от структуры и функционального состояния мембран. Мембрана кардиоцитив имеет ряд особенностей. Как и в других возбудимых тканях, ее интегративные белки-каналы в случае открытия ворот пропускают по градиенту концентрации Na + и Кроме того, здесь есть каналы, которые пропускают Са2 +, Сl-. Эти предварительные каналы отличаются рядом особенностей. Так, Na + проходит не только через «свои», быстрые, каналы, но вместе с Са2 + - через так называемые медленные каналы. Кроме Са2 + -, Na +-канала, Са2 + проникает внутрь и по собственному Са2 +-канала. В мембране содержится не менее двух К +-каналов. Соответствующую функцию в структурах проводящей системы выполняют и Cl - каналы.
В мембране кардиомиоцитов также много белков, которые выполняют функцию ионных насосов. Плотность Na + -, К +-насосов более чем в 100 раз выше, чем плотность
каналов для этих же ионов. их функциональная активность и определяет концентрацию указанных ионов в клетке, таким образом влияя на величину потенциала покоя. Есть здесь большое количество Са2 +-насосов. Плотность насосов, их функциональная активность не постоянны, поскольку регулируются соответствующими системами.
Быстрые Na +-каналы есть и в сократительной миокарде. их кинетика целом аналогична таковой других возбудимых тканей: нервной и поперечно мышечной.
Медленные Са2 +-каналы расположены как в сократительной миокарде, так и в элементах проводящей системы. их кинетика в чем-то напоминает кинетику быстрых Na +-каналов, за исключением скорости: их активационные и инактивацийни ворота медленно открываются и закрываются, а потому медленно развивается деполяризация и восстанавливается возбудимость. При заряде мембраны 0 мВ медленные каналы полностью теряют активность. Пороговый потенциал медленных каналов находится на высоком уровне, чем потенциал быстрых: в медленных каналов он составляет 35-40 мВ, а в быстрых - 55 мВ.
В мембране кардиомиоцитов есть несколько видов К +-каналов. Из них значительные быстрый и медленный. Быстрый канал активизируется
при деполяризации мембраны (потешииалзалежний) до -35 МВГ медленный - при повышении в цитоплазме концентрации свободного кальция (кальцийзависи).
Сl-проходит через каналы пассивно. Низкое содержание в клетке Сl-обеспечивается их выталкиванием наружу отрицательно заряженными ионами в состоянии ВС. При возникновении ПД внутри клетки увеличивается содержание положительных ионов, что приводит к попаданию Сl-в клетку. Поэтому внутриклеточное содержание Сl-зависит от частоты и длительности ПД. Сl-может шунтировать внутриклеточный потенциал, влияя на уровень ПС. Если внеклеточный Сl-заменить менее проницаемой отрицательным ионом, то будет наблюдаться гиперполяризация мембраны.