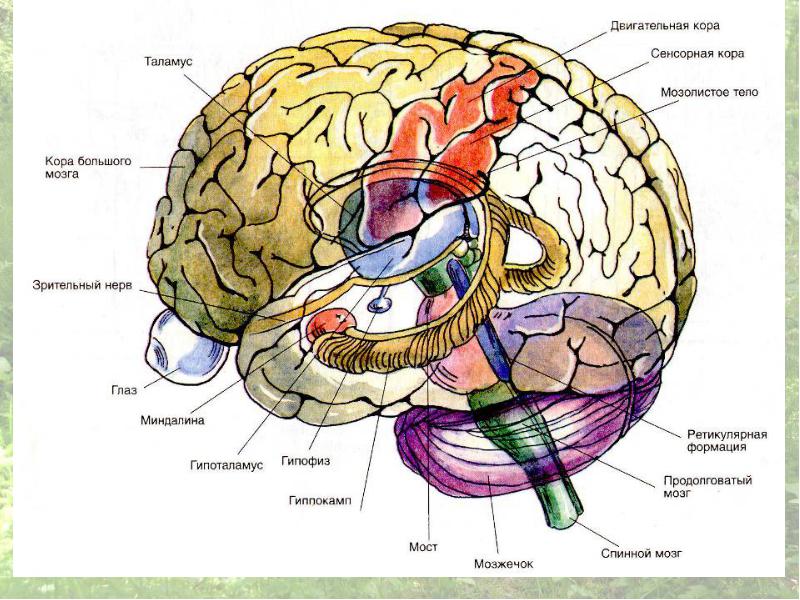
 До этого отдела ЦНС относятся продолговатый мозг и мост головного мозга. Основные функции заднего мозга в регуляции двигательных функций:
До этого отдела ЦНС относятся продолговатый мозг и мост головного мозга. Основные функции заднего мозга в регуляции двигательных функций:
1) анализ сенсорной информации, поступающей в ядра заднего мозга от рецепторов кожи и мышц головы и эфферентная иннервация скелетных мышц головы;
2) анализ сенсорной информации, поступающей от вестибулярных рецепторов, проприорецепторов мышц шеи, туловища, конечностей, и рефлекторная регуляция тонуса мышц тела, которая обеспечивает определенную позу (тонические рефлексы позы);
3) проводниковая функция - прохождение нервных импульсов, идущих в вышележащие отделы ЦНС и от них до скелетных мышц.
Сенсорная информация поступает от рецепторов кожи лица, слизистых оболочек носа, зубов, надкостница костей черепа, проприорецепторов глазодвигательных мышц, вкусовых рецепторов, рецепторов вестибулярного аппарата и слуха, глотки, рецепторов многих внутренних органов.
Эфферентная иннервация обеспечивает функцию жевательных, глазодвигательных, мимических мышц, мышц языка, глотки, гортани, шеи. Эти функции обеспечиваются нервами, еще сохраняют способность к сегментарной иннервации, но она выражена менее отчетливо, чем в спинном мозге.
Очень большую роль играет задний мозг в поддержании позы антигравитации (обеспечивается тонусом мышц) и перераспределении тонуса при изменении положения головы. Жаба, в которой перерезан мозг между продолговатым и спинным (так называемая спинальная животное), не может поддерживать тонус скелетных мышц на таком уровне,
которого было бы достаточно для создания ПОЗЫ антигравитации. Такое животное лежит распластанный на горизонтальной поверхности.
Жаба, в которой перерезан мозг выше заднего, имеет позу, которая позволяет преодолеть силу гравитации Земли. Для создания такой позы важное значение имеет тонус мышц-екстензорив. Этот тонус имеет рефлекторную природу, ибо исчезает после перерезания задних корешков нервов, входящие в спинной или продолговатый мозг. Центрами регуляции есть три двигательные ядра заднего мозга: 1) вестибулярное ядро, 2) ядра ретикулярной формации варолиевого моста, 3) ядра ретикулярной формации продолговатого мозга.
С участием этих центров реализуются тонические рефлексы позы, среди которых различают вестибулярные статические и шейные.
Вестибулярные статические рефлексы начинаются с рецепторов преддверия - сферического и эллиптического мешочков. Они были подробно исследованы голландским физиологом R. Magnus (1924).
Существует несколько видов вестибулярных статических рефлексов:
1) при подъеме головы повышается тонус мышц-екстензорив передних конечностей и снижается тонус этих мышц задних конечностей;
2) при наклоне головы вниз наблюдается противоположный эффект;
3) при наклоне головы в сторону повышается тонус мышц-екстензорив с той стороны, куда наклонена голова.
Значение этих рефлексов заключается в том, что происходит перераспределение тонуса мышц при изменении положения головы, и это обеспечивает поддержание равновесия. Волокна вестибулоспинального пути идут своим боком часто моносинаптических к а-и у-мотонейронов передних рогов спинного мозга. Под влиянием этих волокон возбуждаются а-и у-мотонейроны екстензорив и тормозятся мотонейроны Флексор. При нарушении вестибулярного аппарата или вестибулярных ядер наблюдаются существенные изменения в регуляции тонуса скелетных мышц, а в связи с этим нарушения и в локомоциях.
Шейные тонические рефлексы дублируют вестибулярные статические рефлексы. Эти рефлексы начинаются с проприорецепторов мышц шеи. Импульсы поступают в двигательные центры продолговатого мозга, а оттуда - в мотонейронов спинного мозга. Эти рефлексы влияют не только на мышцы конечностей, но и на глазные мышцы (обеспечивается компенсаторная установка глазных яблок при изменении положения головы). Отклонение головы сопровождается движением глазных яблок в противоположном направлении. Таким образом сохраняется правильная зрительная ориентация. В чистом виде шейные рефлексы можно наблюдать после разрушения вестибулярного аппарата.
Отключить шейные рефлексы можно с помощью перерезания задних корешков соответствующих сегментов спинного мозга или наложение гипсовой повязки на шею, которая делает неподвижными шейные мышцы.
Влияние ретикулярной формации ствола головного мозга на деятельность двигательных центров спинного был открыт И. М. Сеченова в 1862 г. Механизм торможения рефлексов спинного мозга можно объяснить постсинаптическим торможением мотонейронов, влияние на промежуточные интернейроны или на терминале афферентных волокон, входящих в спинной мозг. Сейчас известно, что локальное раздражение гигантоклеточной ядра ретикулярной формации продолговатого мозга приводит к торможению изгибаемых и разгибательных рефлексов спинного мозга.
Кроме тормозных влияний другие участки ретикулярной формации (латеральные зоны) могут облегчать спинальные рефлексы. При этом снижается порог и сокращается латентный период этих рефлексов.
Влияние ретикулярной формации осуществляется через два ретикулоспинальному пути - медиальный и латеральный. Медиальный путь начинается от варолиева моста и, перекрещиваясь, идет к сегментов спинного мозга. При этом возбуждаются мотонейроны екстензорив и тормозятся мотонейроны Флексор. Волокна этого пути проходят в передних отделах спинного мозга рядом с вестибулоспинальним путем и вызывают подобный эффект.
Ретикулярная формация продолговатого мозга действует противоположным образом. Она возбуждает а-и у-мотонейроны Флексор и тормозит мотонейроны екстензорив. Эти влияния реализуются через латеральный ретикулоспинальному пути, часть волокон которого идет по своей стороне, а часть переходит на противоположную сторону.
Двигательные центры ретикулярной формации ствола головного мозга получают многочисленные коллатерали от афферентных путей, идущих в этот отдел ЦНС, а также от эфферентных путей моторной коры, базальных ганглиев, лимбической системы, гипоталамуса.