 Рецепторы, которые участвуют в рефлекторной регуляции дыхания, € в легких, сосудах, головном мозге. По механизму возбуждения они хеморецепторами и механорецепторами.
Рецепторы, которые участвуют в рефлекторной регуляции дыхания, € в легких, сосудах, головном мозге. По механизму возбуждения они хеморецепторами и механорецепторами.
На вентральной поверхности продолговатого мозга у выхода IX и X пар черепных нервов на глубине 200-400 мкм расположены центральные хеморецепторы. Присутствие их можно объяснить необходимостью контроля за поставками 02 мозга, поскольку
при недостатке кислорода быстро погибают клетки ЦНС Ведущим фактором раздражения этих рецепторов является концентрация Н +. Центральные хеморецепторы омываются межклеточной жидкостью, состав которой зависит от метаболизма нейронов и местного кровотока. Кроме этого, состав межклеточной жидкости во многом зависит от состава спинномозговой жидкости.
Спинномозговая жидкость (СМР) отделена от крови ГЭБ. Структуры, его образующих, слабопро-
никнет для Н + и НСО3-»но хорошо пропускают нейтральный С02-Вследствие этого при повышении в крови содержания СОГ он диффундирует в СМР. Это приводит к образованию в ней неустойчивой угольной кислоты, продукты которой стимулируют хеморецепторы. Нужно учитывать, что в норме рН СМР ниже, чем рН крови, - 7,32. Кроме этого, в связи с уменьшением содержания белков буферная емкость СМР также ниже, чем крови. Поэтому при повышении уровня РCO2 в СМР рН изменяется быстрее.
Центральные хеморецепторы оказывают большое влияние на дыхательный центр. Они стимулируют инспираторная и экспираторная нейроны, усиливая как вдох, так и выдох. Поэтому, например, при снижении рН СМР лишь на 0,01 вентиляция легких увеличивается на 4 л / мин.
Периферические хеморецепторы находятся в каротидных тельцах, которые расположены в области бифуркации общих сонных артерий, и в аортальных тельцах, которые есть на верхней и нижней поверхностях дуги аорты. Наибольшее значение для регуляции дыхания имеют каротидные тельца, которые контролируют газовый состав крови, поступающей к мозгу.
Уникальной особенностью рецепторных клеток каротидного синуса является высокая чувствительность к изменениям Ра. При этом рецепторы реагируют на отклонения параметров Раог в очень широких пределах: от 100 до 20 мм рт. ст., и меньше. Чем ниже РаО2 в крови, которая омывает рецепторы, тем больше частота импульсов, идущих от них по нервам Геринга. В основе рецепции лежит собственно интенсивное кровоснабжение тельца - до 20 мл (мин-г). В связи с тем, что 02 в них используется мало, градиент АВРо2 невелик. Поэтому рецепторы реагируют на уровень Рог артериальной, а не венозной крови. Полагают, что механизм раздражения рецепторных клеток при недостатке O2 связан с их собственным метаболизмом, где при малейшем снижении уровня Ро, появляются недоокисленные продукты обмена.
Импульсация от каротидных рецепторов достигает нейронов продолговатого мозга и задерживает вдох, вследствие чего углубляется дыхание. Рефлексы, которые приводят к изменению активности дыхания, возникающие при падении РаО2 ниже 100 мм рт. ст. При этом изменения дыхания при раздражении каротидных хеморецепторов наступают очень быстро. их можно обнаружить даже в течение одного дыхательного цикла при относительно незначительных колебаниях концентрации газов в крови. Раздражаются эти рецепторы также при снижении рН или повышении расой. Гипоксия и гиперкапния взаимно усиливают импульсацию от этих рецепторов.
Меньшее значение для регуляции дыхания имеют аортальные хеморецепторы, которые играют заметную роль в регулировании кровообращения.
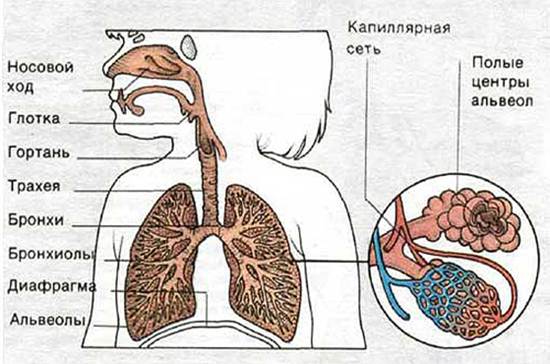
Рецепторы легких и воздухоносных путей. Эти рецепторы относятся к механо-и хеморецепторов. В гладких мышцах воздухоносных путей, начиная от трахеи и заканчивая бронхами, содержатся рецепторы растяжения легких. В каждой из легких является до 1000 рецепторов.
Выделяют несколько типов рецепторов, которые реагируют на растяжение легких. Около половины рецепторов раздражаются только при глубоком вдиси. Это пороговые рецепторы. Низькопорогови рецепторы раздражаются и при малом объеме легких, т.е. во время как вдоха, так и выдоха. Во время выдоха частота импульсации от этих рецепторов возрастает.
Механизм раздражения рецепторов легких заключается в том, что мелкие бронхи растягиваются за счет их эластичности, которая зависит от степени расширения альвеол; что оно больше, тем сильнее растяжение структурно связанных с ними воздухоносных путей. Большие воздухоносные пути структурно связаны с легочной тканью и раздражаются из-за «негативности давления» в плевральной щели.
Рецепторы растяжения относятся к таким, которые мало способны к адаптации, и при длительной задержке вдоха частота импульсов от легких уменьшается медленно. Чувствительность этих рецепторов не постоянная. Например, при бронхиальной астме за счет спазма бронхиол возбудимость рецепторов возрастает. Поэтому рефлекс появляется при меньшем растяжении легких. Состав воздуха, содержащегося в легких, также влияет на чувствительность рецепторов. При увеличении уровня С02 в воздухоносных путях импульсация с рецепторов растяжения уменьшается.
Большинство афферентных импульсов от рецепторов растяжения легких направляется в дорсального ядра бульбарного отдела дыхательного центра и активизирует И (5-нейроны. В свою очередь эти нейроны, тормозя активность Иа-нейронов, останавливают вдох. Но такие реакции наблюдаются только при высокой частоте импульсов, которая достигается на высоте вдоха. При низкой частоте рецепторы растяжения, наоборот, продолжают вдох и сокращают выдох. Думают, что относительно редкие разряды, которые поступают во время выдоха от рецепторов растяжения, способствующие наступлению вдоха.
У человека рефлексы, связанные с раздражением механорецепторов легких (рефлексы Геринга-Брейер), большого значения не имеют, они только предотвращают чрезмерное растяжение легких при вдыхании более 1,5 л воздуха.
Иритантные рецепторы расположены в эпителиальном и субэпителиального слоях воздухоносных путей. Особенно много их в области корней легких. Импульсы от этих рецепторов идут по миелиновых волокнах блуждающих нервов. Иритантни рецепторы обладают одновременно свойствами механо-и хеморецепторов. Они быстро адаптируются. Раздражителями этих рецепторов также едкие газы, холодный воздух, пыль, табачный дым, биологически активные вещества, образующиеся в легких (например, гистамин).
Раздражение иритантних рецепторов сопровождается неприятным ощущением - жжением, кашлем и др.. Импульсы из этих рецепторов, которые поступают за счет более раннего вдохе, сокращают выдох. Вероятно, «морковь» (в среднем 3 раза за 1 год), которые возникают при спокойном дыхании, также обусловлены рефлексами с иритантних рецепторов. До того как возникают «морковь», нарушается равномерность вентиляции легких. Это приводит к раздражению иритантних рецепторов и углубляется один из вдохов, вследствие чего расширяются отделы легких, которые до этого спасшиеся. Раздражение иритантних рецепторов через блуждающий нерв может привести к сокращению гладких мышц бронхов. Этот рефлекс лежит в основе бронхоспазма при возбуждении рецепторов гистамином, который образуется при бронхиальной астме. Физиологическое значение указанного рефлекса состоит в том, что при вдыхании токсических веществ изменяется просвет бронхов, снижаются вентиляция альвеол и газообмен между дыхательными путями и альвеолами. Благодаря этому в альвеолы и кровь попадает меньше токсичных веществ.
J-рецепторы, или юкстамедулярни рецепторы, называются так потому, расположенные в стенках альвеол у капилляров. Раздражаются они при поступлении биологически активных веществ в малый круг кровообращения, а также при увеличении объема интерстициальной жидкости легочной ткани. Импульсы от них идут в продолговатый мозг по немиелинизованих волокнам блуждающего нерва. В норме J-рецепторы находятся в состоянии слабого тонического возбуждения. Усиление импульсации приводит к частому поверхностного дыхания. Роль этих рецепторов в регуляции дыхания неизвестна. Возможно, они вместе с иритантнимы рецепторами вызывают одышку при набухании легких.
На регулирование дыхания влияют импульсы еще от нескольких типов рецепторов.
Рецепторы плевры относятся к механорецепторов. Они играют определенную роль в изменении характера дыхания при нарушении свойств плевры. При этом возникает ощущение боли, главным образом ииовьязане с раздражением париетальной листка плевры.
Рецепторы верхних дыхательных путей реагируют на механические и химические раздражители. Они подобны иритантних рецепторов. их раздражение вызывает чихание, кашель и сужение бронхов.
Рецепторы дыхательных мышц. Мышечные веретена дыхательных мышц (межреберных мышц и мышц стенки живота) возбуждаются как при растяжении мышцы, так и по принципу гема-петли. Рефлекторные дуги из этих рецепторов замыкаются на уровне соответствующих сегментов спинного мозга. Физиологическое значение этих рефлексов заключается в том, что при затруднении дыхательных движений автоматически усиливается сила сокращения мышц. Сопротивление дыханию увеличивается, например, при уменьшении эластичности легких, бронхоспазме, отека «слизистой оболочки внешнем сопротивления расширению грудной клетки. В обычных условиях проприорецепторы дыхательных мышц значительной роли не играют. Но их влияние легко обнаружить при интенсивном сжатии грудной клетки, при котором они включают вдох. В диафрагме содержится очень мало рецепторов (10-30), и они не играют существенной роли в регулировании дыхания.
Рецепторы суставов и «недыхательном» скелетных мышц играют определенную роль в поддержании рефлекторной одышки при выполнении физической работы. Импульсы от них достигают бульбарного центра ди-
зияння.
Раздражение болевых и температурных рецепторов может рефлекторно влиять на характер дыхания. Чаще наблюдается начальная задержка дыхания с последующей одышкой. Гипервентиляция может возникнуть и при раздражении температурных рецепторов кожи. Вследствие этого возрастает частота дыхания при уменьшении его глубины. Это способствует увеличению вентиляции легочного пространства и выделению избытка тепла.