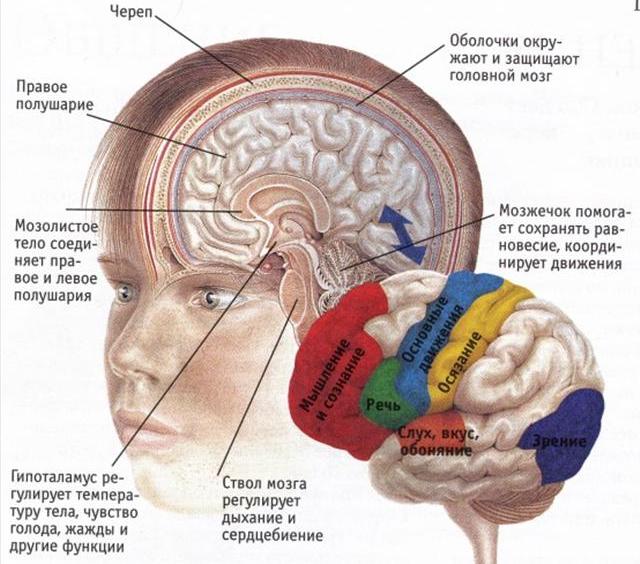
 Кора большого мозга - филогенетически молодая структура мозга. В процессе эволюции млекопитающих особенно быстро развивалась новая кора, толщина достигла 1-2 мм, а общая ее поверхность у человека составляет около 2200 см2. В состав коры большого мозга входят нейроны, которые в разных ее участках имеют свои особенности. Различают свыше 50 цитоархитектурних полей. Многие из них участвуют в регуляции двигательных функций.
Кора большого мозга - филогенетически молодая структура мозга. В процессе эволюции млекопитающих особенно быстро развивалась новая кора, толщина достигла 1-2 мм, а общая ее поверхность у человека составляет около 2200 см2. В состав коры большого мозга входят нейроны, которые в разных ее участках имеют свои особенности. Различают свыше 50 цитоархитектурних полей. Многие из них участвуют в регуляции двигательных функций.
Сенсорные зоны коры большого мозга тесно связаны с моторными зонами, которые лежат перед центральной бороздой, занимая примерно заднюю часть лобной доли. В моторной коре различают: первичную, премоторной и дополнительной моторной области.
Первичная моторная кора находится перед центральной бороздой (поле 4). Как и первая соматосенсорная зона, участок имеет топической организации. Раздражение электрическим током верхней части участка у животных или у людей во время нейрохирургических операций сопровождается сокращением мышц нижней части тела на противоположной стороне. Более 50% поверхности этого участка у человека - это представительство рук и языка, что свидетельствует о развитии этих функций у человека.
Премоторной зона лежит впереди первичной моторной (поле 6) и в глубине сильвиевой борозды. Большинство импульсов из этой зоны вызывает модели движения, включаются группы мышц, которые выполняют определенные функции. Импульсы идут в первичную моторную кору, в базальных ядер, а затем снова в моторную зону через таламус. Эта зона вместе с базальными ядрами, таламусом, первичной моторной корой управляет многими сложными движениями.
Дополнительная моторная участок расположен перед и над премоторной зоной. Чтобы "сократились мышцы, контролируемые этой зоны, нужно применить электрический ток значительно большей мощности. Эффект раздражения часто проявляется с обеих сторон тела. Функция рассматриваемой области состоит в том, что она вместе с премоторной участком создает условия для обеспечения позы и движений, которые реализуются уже первичной моторной корой. Нейроны коры моторной зоны получают многочисленные импульсы волокнами, идущими от сенсомоторной области париетальной коры, зрительной и слуховой участков, моторной коры противоположной стороны, таламуса, а также от базальных ядер и других структур.
Кортико-спинальный (пирамидный) путь начинается от нейронов моторной коры, проходит между хвостатым ядром и скорлупой (внутренняя капсула). В продолговатом мозге большинство волокон (80%) переходит на противоположную сторону - в латеральный пирамидный путь, а часть (20%)-на противоположную сторону ниже (в шейном и грудном отделах спинного мозга), т.е. в вентральный пирамидный путь.
В пирамидному пути содержится около 1 млн волокон. Лишь незначительная часть (около 3%) начинается от гигантских пирамидных клеток Беца. Коллатерали от волокон пирамидного пути идут в соседние участки коры головного мозга и вызывают торможение (таким образом предотвращают иррадиации возбуждения). Большинство волокон латерального пирамидного пути заканчивается на интернейронах спинного мозга, часть - на сенсорных нейронах задних рогов или собственно на мотонейронах.
Роль пирамидного пути в осуществлении двигательной функции заключается в регуляции тонких и точных движений, особенно дистальных частей конечностей, т.е. пальцев. При нарушении кровоснабжения моторной коры (кровоизлияние, тромбоз, травма, опухоль) наблюдается паралич мышц на противоположной стороне тела. Значительное количество волокон от моторной коры не идет непосредственно к спинному мозгу пирамидным путем, а переключается на различных ядрах - базальных, красном, вестибулярных, ядрах ретикулярной формации и др..
Раньше широко пользовались термином «экстрапирамидная система». Теперь его употребляют реже, потому пути, входящих в его состав, имеют различное функциональное назначение. Моторная кора тесно связана с красным ядром. Это кортико-руброспинальный путь. Аксоны от коры заканчиваются синапсами в нижней части красного ядра у больших клеток. В красном ядре тоже являются топические представительство всех мышц тела. Отсюда начинается руброспинальный путь, переходит на противоположную сторону. Его волокна контактируют с интернейронов спинного мозга, а некоторые - прямо с мотонейронами, которые контролируют функцию преимущественно дистальных мышц тела.
Кортико-спинальный и руброспинальный пути называют еще латеральной моторной системой, а вестибулоспинальний и ретикулоспинальному - медиальной моторной системой.
Колончастый организация зон коРй состоит из вертикально расположенных групп нескольких тысяч нейронов, которые посылают импульсы к мышце или мышц-синергистов. Эти группы нейронов называют колонками.
Каждая колонка содержит пирамидные клетки в V слое, имеет вход во II слой через клетки IV слоя., С другими участками коры она связывается через клетки VI слоя. Каждая колонка получает информацию из многих источников. Одна пирамидная клетка не может вызвать возбуждение мышцы, для этого нужно, чтобы в состояние возбуждения пришли 50-100 пирамидных клеток. Различают динамические и статические сигналы этих клеток. Динамические сигнал-импульсы, в генерируемых динамическими нейронами, их частота больше и приводит к сокращению мышцы. Статические сигналы - импульсы статических нейронов, их частота меньше, но действие длительнее. Они лишь пидгримують сокращения мышцы.
Таким образом, а-мотонейроны переднего рога спинного мозга анализируют и синтезируют информацию, поступающую к ним от сенсорных нейронов своего и соседних сегментов, от пирамидного, руброспинальный, вестибулоспинального, ретикулоспинальному и других путей через интернейроны или непосредственно.