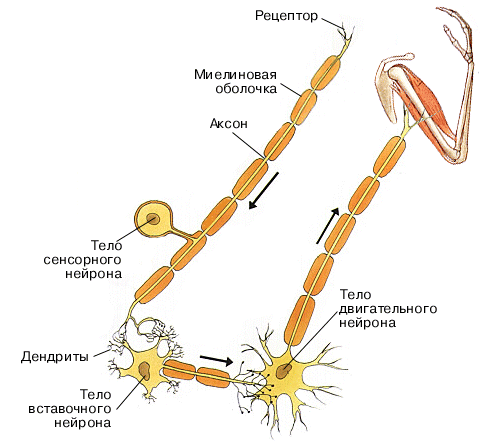
 Взаимодействие нейронов и нервных процессов (возбуждения и торможения) в ЦНС, которая обеспечивает ее согласованную деятельность, носит название координации (согласования).
Взаимодействие нейронов и нервных процессов (возбуждения и торможения) в ЦНС, которая обеспечивает ее согласованную деятельность, носит название координации (согласования).
Благодаря координации возможно точное выполнение в данный момент различных сложных рефлекторных актов, к которым относятся двигательные, секреторные, сосудистые и другие компоненты. Координация происходит во всех отделах ЦНС, в любом нервном центре.
Основные принципы координации рефлексов: реципрокные (объединенная) иннервация, общий конечный путь; принципы обратной связи и доминанты. В координации рефлексов участвуют также процессы иррадиации возбуждения, конвергенции и дивергенции, явления облегчения и окклюзии ".
Реципрокные иннервация. В основе этого явления лежит представление о том, что возбуждение центра одной группы мышц (например, мотонейронного «пула» мышц-сгибателей) сопровождается этой стороне реципрокным (совмещенным) торможением центров мышц-антагонистов (разгибателей). Одновременно на противоположной стороне (в симметричных конечностях) возбуждаются центры разгибателей и тормозятся мотонейроны мышц-сгибателей. Конечно, только при этом условии возможен точное движение сгибания или разгибания. Реципрокные взаимоотношения между группами мотонейронов и вставочных нейронов лежат в основе так называемых ритмических рефлексов, которые характеризуются правильным чередованием противоположных по своему функциональному значению мышечных сокращений, например-сгибание и разгибание. Примером таких ритмических рефлексов у млекопитающих могут быть рефлексы почесывание и шаговый, возникающие в целом при раздражении рецепторных полей длинным и слабым раздражением.
Механизм реципрокных взаимоотношений заключается в том, что афферентные нервные волокна (например, группы Аа от мышечных веретен) дают в спинном мозге ветвления. Одни из ветвей образуют возбуждающие синапсы на мотонейронах мышц-сгибателей, а другие с помощью вставочных нейронов - тормозные синапсы на мотонейронах мышц-разгибателей. Поэтому раздражение афферентных нерва вызывает одновременное возбуждение мотонейронов мышц-сгибателей и торможения мотонейронов мышц-разгибателей. Реципрокное торможение имеет короткую рефлекторную дугу среди известных.
Однако было бы ошибкой считать, что между центрами существуют навсегда фиксированные антагонистические отношения. Они имеют динамический характер, могут определяться различными условиями. Нет также оснований ограничивать реципрокные отношения участком иннервации анатомических анастомозов. В такие же реципрокные отношения могут вступать и другие, отделены друг от друга центры, когда они регулируют определенные состояния какого-либо органа или системы »
Подобные реципрокные взаимоотношения наблюдаются между центрами вдоха и выдоха, голода и насыщения, дыхания и глотания подобное. Однако реципрокные отношения между различными центрами не являются постоянными, они могут меняться (прекращаться) под влиянием коры и подкорковых структур большого мозга.
Принцип общего конечного пути. Ch. S. Sherrington (1906) ввел-понятие о мотонейрон как общий конечный путь двигательной системы, то есть путь, которым осуществляется переработка и интеграции всех многочисленных и разнородных возбуждений, одновременно поступающих к нему, в единое (аксонного) возбуждение, которое идет в эффектора (м мышцы). Структурной основой общего конечного пути является значительное превышение в большинстве нейронов ЦНС количества афферентных «входов» над эфферентным «выходом»-наличие своеобразной «воронки», расширенная часть которого символизирует число афферентных волокон, разветвления которых конвертируются в мо «тонейрони, а суженная часть - общий конечный путь, которым является аксон мотонейрона.
Поскольку нейрон интегрирует как возбуждающие, так и тормозные процессы, ПД возникает лишь в том случае, когда преобладают возбуждающие явления, т.е. когда ВПСП в области аксонного бугорка результате суммации достигает критического уровня. Иначе говоря, эти влияния конкурируют за овладение общим конечным путем. Результат этой «борьбы» за общий конечный путь зависит от силы действующих раздражений, их биологического значения в данной ситуации, от состояния нервных центров, особенно доминантного. Рефлексы, дуги которых имеют общий конечный путь, могут взаимно усиливаться (в случае так называемых союзных рефлексов) или ослабляться (в случае антагонистических рефлексов).
Принцип доминанты. Известный русский физиолог А. А. Ухтомский (1875-1942) сформулировал принцип доминанты, который является одним из основных принципов деятельности ЦНС;
Доминанта (от лат. Dominare - властвовать)-это господствующие в ЦНС очаги возбуждения, которые изменяют и «подчиняют» себе в данный момент деятельность других центров.
Доминанта формируется под влиянием различных адекватных стимулов внешней и внутренней среды организма (истощение питательных веществ, накопление в крови половых гормонов и др.). Все эти стимулы повышают возбудимость соответствующих нервных центров (пищевого, полового и т.д.). За счет суммации с посторонними (индифферентными) стимулами разной модальности в данном центре формируется стойкое и длительное возбуждение, сопряженное с торможением других, связанных с ним, центров. Так возникают пищевая, половая, оборонная и другие доминанты, которые в значительной мере определяют осуществления именно данного рефлекса, поведение человека и животных в данный момент.
Можно любой очаг возбуждения, возникшего в ЦНС, рассматривать как доминантный? Какие характерные черты доминанты? Доминантный очаг возбуждения (любой центр) характеризуется следующими свойствами: 1) Повышение возбудимости, 2) устойчивость возбуждения, 3) способность к суммации возбуждения (накапливать возбуждения за счет сторонних раздражений) 4) способность тормозить другие нервные центры и системы рефлексов, 5 ) способность длительное время удерживать возбуждения по окончании раздражение, которое его обусловило (иннерция доминанты).
"Примерами доминанты могут быть следующие наблюдения. Раздражение у животных некоторых участков коры большого мозга, которое обычно сопровождается сгибанием конечностей, во время акта дефекации или мочеиспускания такой реакции не вызывает. Наоборот, происходит усиление этих рефлекторных актов. Это обусловлено тем, что в данный момент центры дефекации или мочеиспускания находятся в состоянии доминантного возбуждения, который «притягивает» к себе посторонние возбуждения и оказывает сочетанное торможение других нервных центров.
Доминантный состояние может возникать и под влиянием гормонов, которые повышают возбудимость соответствующих центров. Например, у лягушек-самцов в весенний период заметно усиливается половой (охватывающий) рефлекс под влиянием посторонних (для полового центра) раздражений (например, при щипки кавычки). В то же время возникает защитного изгибающего рефлекса этой конечности.
Явления, которые напоминают доминанту, приходится наблюдать и в клинической практике, когда источники возбуждения, возникшие в ЦНС, подобно магниту, «притягивают» к себе различные посторонние раздражения, усиливаясь за их счет. Так, при язвенной болезни желудка и двенадцатиперстной кишки в стадии доминантного возбуждения находится пищевой центр, при гипертонической болезни - сосудодвигательный, вследствие чего различные возбуждение (особенно эмоциональные) приводят к повышению желудочной секреции или уровня системного артериального давления. После травмы нервных стволов нередко в состоянии доминантного возбуждения находятся центры болевой чувствительности в таламокортикальных структурах МОАК, вследствие чего усиливается боль в раненой конечности (каузалгия) при различных посторонних раздражениях (сильном зуде, притрагивании к любой части тела и т.п.).
Прекращаться доминанта может разными путями: 1) вследствие устранения раздражения или удовлетворения потребностей (биологической, социальной), на основании которых сформировалась доминанта, 2) при возникновении в ЦНС сильной доминанты, тормозящей ту, которая является, 3) вследствие развития в доминантном центре вторичного торможения.
Благодаря доминанте в ЦНС создаются не стабильны, а динамические координационные отношения между нервными центрами и системами рефлексов. Доминанта играет важную роль в психической деятельности человека и животных, в значительной мере определяет поведенческие акты в определенный момент.
Облегчение рефлексов. Если раздражение двух рецепторных полей настолько слабо, что каждое из них (хотя и возбуждает эфферентные волокна) все же не вызывает разряд нейронов и рефлекторный ответ, то при одновременном раздражении может возникнуть четкая рефлекторная реакция, которая превысит по интенсивности ответа на каждый из отдельных раздражителей. Это явление получило название облегчения рефлексов. В его основе лежит суммация ВПСП, которая происходит в нейронах подпороговых зоны нервного центра и влечет возникновение у них ПД.
Окклюзия рефлекса. При одновременном сильном раздражении двух афферентов рефлекторный ответ может быть значительно меньше, чем сумма таких ответов на каждый из раздражений, или одинаковой с ней. Это явление уменьшения ответа получило название окклюзии (закупорки) и ничего общего с торможением нет. В основе этого явления лежит общность (перекрытия) зон разряда двух или нескольких афферентных входов нервного центра. Так, при раздражении источника А розряджуется 11 нейронов, а при раздражении источника Б - 9 нейронов, 5 из которых уже втянулись в импульсную реакцию при раздражении источника А. Поэтому общий
не число нейронов, розряджуються, будет при одновременном раздражении двух афферентных входов меньше арифметической суммы нейронов, которые генерируют ПД на каждое из раздражений отдельно. Так, нанесение раздражений на два источника (А и Б) вызывает импульсную реакцию только у 16 нейронов, а не в 20, как это должно быть при простой суммации активности нейронов, разряжаются на каждое из раздражений (А или Б).
Принцип обратной связи. Изучением роли «обратной связи» или «обратной афферентации» в механизмах координации рефлекторной деятельности занимались П. К. Анохин (1945), Н. А. Берн-штейн (1966), Ch. S. Cherrington (1906).
Суть принципа обратной связи заключается в том, что при совершении любого рефлекторного акта процесс не ограничивается рабочим эффектом (например, мышечным движением), а сопровождается возбуждением рецепторов рабочего органа (мышцы и т.п.), от которых информация о последствиях действия поступает афферентными проводниками в ЦНС. Там она сопоставляется, сравнивается с запрограммированным, ожидаемым результатом. Поэтому афферентные сигналы, поступающие в ЦНС от рецепторов исполнительных органов, принадлежащих к той или рефлекторной дуги, получившие название обратной, или вторичной, афферентации (в отличие от тех сигналов, которые первично вызывали рефлекторный акт).
Роль обратной афферентации в механизмах координации очень велика. Благодаря обратной связи интенсивность и последовательность включения различных групп нейронов становятся строго согласованными, координированными с результатом действия (мышечное движение, изменение уровня кровяного давления, выделение секрета т.п.), т.е. осуществляется контроль за эффективностью реакции. Тем самым создается возможность для внесения поправок, коррекции осуществления рефлекторного акта. Поэтому неудивительно, что при повреждении мышечной чувствительности движения, особенно при ходьбе, становятся неточными, некоординированными, порывистыми. Это происходит вследствие потери контроля над движениями со стороны ЦНС, что обусловлено повреждением каналов обратной афферентации.